
1.2 GRAS转录因子家族
GRAS转录因子家族是一类近来才被研究的基因家族[1, 2, 5]广泛存于许多高等植物中,例如拟南芥、水稻、豌豆、大豆和大麦等植物。GRAS家族的名字起源于三个最初发现分离出来的成员GAI [1, 2, 5][GA (GIBBERELLIC ACID)-INSENSITIVE],RGA(REPRESSOR OF GAI)和SCR (SCARECROW)。根据研究可以对GRAS家族进行分:DELLA、HAM、LISCL、PAT1、LS、SCR、SHR 和SCL3共8类。GRAS转录因子蛋白,在N端有特异性并且拥有高度的可变性[1, 2, 5],而C-端是稳定的保守结构拥有很高的同源性(图一)。两个亮氨酸丰富的区域从侧面夹击一VHIID基序(motif),这是GRAS家族的标志性结构域。VHIID基序处于N端和C端之间,并指出了若干重要的氨基酸。
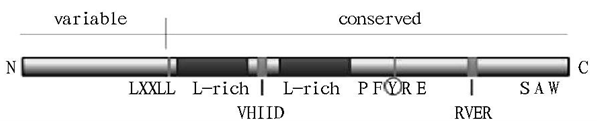
图一:GRAS 转录因子蛋白结构图解 [1, 3, 5]
Fig1: GRAS transcription factor protein structure diagram
1.3 GRAS在植物中的分布
GRAS蛋白家族在许多植物中是常见的,并且是植物中一种特殊的蛋白质家族。当前,我们在拟南芥中发现了33个GRAS转录因子[4],主要包括:SCL、SCR 、SHR、GAI、RGA、RGL和PAT1等。在水稻中发现了60 个GRAS 家族成员[2, 6-8],如MOC1、SLR1和CIGR1 /2等。通过数据库检索番茄中有51GRAS基因家族序列。番茄GARS家族成员在12条染色体上都有分布[9],主要位于染色体两端。此外,还在烟草中分离发现了一个核定位的GRAS 基因家族新成员NtGRAS1,并指出NtGRAS1基因可能作为一个重要的转录调节因子参与植物胁迫反应。有研究指出,葡萄也有GRAS 基因家族[10],同拟南芥GRAS 基因家族一样拥有很高的同源性。运用基因同源性克隆技术从大赖草中克隆了LRC1 基因[6, 7, 11, 12],该基因与调节水稻分蘖的MOC1基因拥有很高的相似性( 达91%) 。此外,在番茄、玉米、矮牵牛、大麦、百脉根、松树、胡杨、佛手、黄瓜、海马齿、苜蓿、胡杨和白桦等很多植物中也发现了GRAS转录因子家族的身影。论文网
1.4 GRAS的功能研究
GRAS基因家族是独特的转录因子家族,在植物中GRAS蛋白数量众多,功能百般,在很多重要的生长发育过程当中起到了至关重要的调控作用。例如:在番茄,水稻中,GRAS 基因家族(LS、OsMOC1)参与了侧生分生组织的发育[6, 7, 12, 13];在矮牵牛花中 ,GRAS 基因家族(PhHAM)参与了茎尖分生组织的形成;在拟南芥中,GRAS基因家族(SHR,SCR,SCL)参与根辐射形成,GRAS基因,GRAS 基因家族(GAI、RGA、RGL)参与了赤霉素的信号转导[2, 5, 15],GRAS 基因家族(SCL13、PAT1、SCL21)对光敏色素信号传导有着影响等。其中DELLA蛋白作为GRAS的分支蛋白对赤霉素(GA)通道具有负调节作用,能够抑制植物的生长发育。通过对DELLA蛋白的降解来调节植物生长发育[4, 12, 14],当DELLA蛋白在GA感知区域被快速降解,则其阻遏的功能消除,植株表现出正常的GA反应。而DELLA蛋白的结构发生变化,抑制蛋白质的降解,表现出不敏感的株矮化表型。假如DELLA蛋白大多数的功能丧失,植株表现出对GA持续反应得细长型表型[8, 10, 15]。SCARECROW作为第一个被鉴定的GRAS家族成员与SHORTROOT 共同调节根的发育过程。SCR对细胞的不对称分裂进行调控,功能缺失的突变植物只能形成单层细胞细胞组织。而与SCR不同SHR在中柱细胞内表达,向外利用单层细胞进入邻近的内皮层细胞进行调控。SCR和SHR共同作用下形成一个对根的正反馈调节机制。GRAS蛋白的PAT1亚家族成员分支中的PAT1,SCL221和SCL13是影响光敏色素信号传导的中间介质[1, 4, 9]。其中SCL13脱离了phyB,独自调控调节光敏色素phyA的信号转而白SCL21可以与PAT1相互作用, 共同作用于phyA的信号转导。另外,还发现GRAS基因家族面对不同的胁迫中有着不同反应应答,通过拟南芥GRAS基因家族在渗透作用下的应答表现以及在干旱胁迫过程当中的应答表现进行剖析, 我们看到了有快要有10个GRAS家族基因在渗透作用和干旱胁迫过程中时有显著上升的现象。Torres-Galea检测拟南芥PAT1分支AtSCL13应答表现时看出, 在盐、干旱、低温等条件下均能诱导表达AtSCL13[2, 5, 15] 。水稻GRAS家族PAT1分支中的2个基因OsCIGR1和OsCIGR2[6] , 和真菌共同培养时都可以被剧烈的诱发表现,表明这2个基因在水稻防御反应的早期有着重要的调控作用。佛手中的PAT1分支基因家族,在低温环境下,其应答更加明显;白菜中的BoGRAS (SCL13)是一类对热环境应答的基因,可以筛选出具有耐热性能的白菜的分子标记。这些结果说明了GRAS家族的分支PAT1基因不仅对光敏色素信号转导有着至关重要的调控作用, 也直接作用植物的抗逆过程。DELLA蛋白也能够提高植物的耐受能力,Achard和Genschik[2, 6] 发现DELLA蛋白可以通过调控活性氧ROS水平来调节植物对胁迫的应答和发育,增强植物的抗性。DELLA[1]蛋白还在GA信号途径和依靠于CBF1的冷信号传导途径交互过程中发挥作用,CBF1利用对DELLA蛋白的诱发累积,从而对植物的生长发育有着负调节。另外, 还有其他的GRAS基因家族对植物的胁迫逆境也有影响, 如烟草GRAS基因(NtGRAS1)在受到C26H36N2O9、双氧水、C7H6O3作用时表现出显著上升的趋势;通过表达胡杨GRAS基因SCL7[2, 6] ,能够明显增强转基因拟南芥的耐盐性和耐旱性。根据研究可以看出,水稻OsGRAS1在干旱、盐及外源ABA的诱发下表现为上调,利用对水稻OsGRAS1基因启动子序列的分析, 发现其含有多个脱水响应元件、ABA响应元件及伤害响应元件,可以看出该基因有可能参与水稻的干旱应答、逆境调控。这些可能是未来GARS基因家族研究的重点[2, 4]。 玉米GRAS基因家族在植物中的进化析(2):http://www.751com.cn/shengwu/lunwen_76314.html